Echidna Development
| Embryology - 26 Apr 2024 |
|---|
| Google Translate - select your language from the list shown below (this will open a new external page) |
|
العربية | català | 中文 | 中國傳統的 | français | Deutsche | עִברִית | हिंदी | bahasa Indonesia | italiano | 日本語 | 한국어 | မြန်မာ | Pilipino | Polskie | português | ਪੰਜਾਬੀ ਦੇ | Română | русский | Español | Swahili | Svensk | ไทย | Türkçe | اردو | ייִדיש | Tiếng Việt These external translations are automated and may not be accurate. (More? About Translations) |
Introduction

The Echidna family consists of 2 major groups the short-beaked in Australia and long-beaked in New Guinea and Indonesia (Irian Jaya). Together with the platypus these are the only 3 surviving genera of the order Monotremata.

The echidna is a unique egg-laying mammal, the embryo is referred too as a "puggle" (not to be confused with the dog breed, produced by mating a Pug with a Beagle) and is not a common animal model of mammalian embryonic development.
The New Guinea long-beaked echidna (Zaglossus bruijni bartoni) is currently on the endangered category (More? Zoo Threatened Species list)
Historic Embryology: 1887 Monotremata and Marsupialia Embryology | 1915 Monotreme Skull | 1932 Spleen | 1939 Growth of the ovarian ovum, maturation, fertilisation and early cleavage | 1939 motor cortex and pyramidal tract | Hill Collection contains echidna embryonic development | Category:Echidna
| Australian Animal: echidna | kangaroo | koala | platypus | possum | Category:Echidna | Category:Kangaroo | Category:Koala | Category:Platypus | Category:Possum | Category:Marsupial | Category:Monotreme | Development Timetable | K12 | |||||
|
| ||||
Some Recent Findings
|
| More recent papers |
|---|
 This table allows an automated computer search of the external PubMed database using the listed "Search term" text link.
More? References | Discussion Page | Journal Searches | 2019 References | 2020 References Search term: Echidna Development | Echidna Embryology |
| Older papers |
|---|
| These papers originally appeared in the Some Recent Findings table, but as that list grew in length have now been shuffled down to this collapsible table.
See also the Discussion Page for other references listed by year and References on this current page.
|
Taxon
Short-beaked Echidna - Tachyglossus aculeatus
Long-beaked Echidna - Zaglossus bruijni
Tachyglossus aculeatus Lineage (full) cellular organisms; Eukaryota; Fungi/Metazoa group; Metazoa; Eumetazoa; Bilateria; Coelomata; Deuterostomia; Chordata; Craniata; Vertebrata; Gnathostomata; Teleostomi; Euteleostomi; Sarcopterygii; Tetrapoda; Amniota; Mammalia; Prototheria; Monotremata; Tachyglossidae; Tachyglossus
Echidna Zaglossus bruijn Lineage (full) cellular organisms; Eukaryota; Fungi/Metazoa group; Metazoa; Eumetazoa; Bilateria; Coelomata; Deuterostomia; Chordata; Craniata; Vertebrata; Gnathostomata; Teleostomi; Euteleostomi; Sarcopterygii; Tetrapoda; Amniota; Mammalia; Prototheria; Monotremata; Tachyglossidae
Development Overview
Gestation is from 22 to 23 days. (based upon 20 observed matings and documenting 30 incidences of egg laying, Rismiller, 1999).
Egg only a single egg is generally laid.
Incubation lasts for approximately 10 days after laying, the hatched embryo (puggle) requires further development.
Embryo after hatching hangs from hairs and succles from a "mammary gland" (mammary hairs) in the pouch for approximately 50 days and continues to develop.


Historic drawings of Echidna embryology (1894).[11]
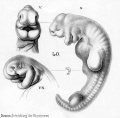
drawing 40
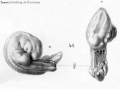
drawing 41
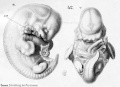
drawing 42
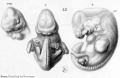
drawing 43

drawing 44
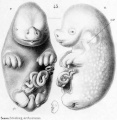
drawing 45

drawing 46
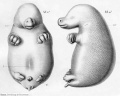
drawing 47
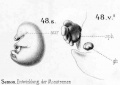
drawing 48
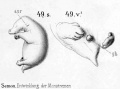
drawing 49
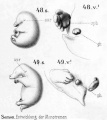
drawing 48-49

drawing 50

drawing 51

drawing 52
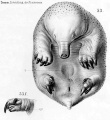
drawing 53















Evolution
The oldest platypus and its bearing on divergence timing of the platypus and echidna clades.[12]
- "Monotremes have left a poor fossil record, and paleontology has been virtually mute during two decades of discussion about molecular clock estimates of the timing of divergence between the platypus and echidna clades. ...Strict molecular clock estimates of the divergence between platypus and echidnas range from 17 to 80 Ma, but Teinolophos (Early Cretaceous fossil) suggests that the two monotreme clades were already distinct in the Early Cretaceous, and that their divergence may predate even the oldest strict molecular estimates by at least 50%."
Hill Embryological Collection
James Peter Hill (1873-1954) University of Edinburgh, Royal College of Science in London, 1892 demonstrator in Sydney, Australia. In 2004 this embryo collection was relocated to the Museum fur Naturkunde, Berlin.
References
- ↑ 1.0 1.1 Morrow G & Nicol SC. (2009). Cool sex? Hibernation and reproduction overlap in the echidna. PLoS ONE , 4, e6070. PMID: 19562080 DOI.
- ↑ Anthwal N, Fenelon JC, Johnston SD, Renfree MB & Tucker AS. (2020). Transient role of the middle ear as a lower jaw support across mammals. Elife , 9, . PMID: 32600529 DOI.
- ↑ Ashwell KWS, Shulruf B & Gurovich Y. (2019). Quantitative Analysis of the Timing of Development of the Cerebellum and Precerebellar Nuclei in Monotremes, Metatherians, Rodents, and Humans. Anat Rec (Hoboken) , , . PMID: 31633884 DOI.
- ↑ Morrow GE, Jones SM & Nicol SC. (2017). Frozen embryos? Torpor during pregnancy in the Tasmanian short-beaked echidna Tachyglossus aculeatus setosus. Gen. Comp. Endocrinol. , 244, 139-145. PMID: 26562301 DOI.
- ↑ Alibardi L & Rogers G. (2015). Observations on fur development in echidna (Monotremata, Mammalia) indicate that spines precede hairs in ontogeny. Anat Rec (Hoboken) , 298, 761-70. PMID: 25367156 DOI.
- ↑ Ashwell KW. (2012). Development of the hypothalamus and pituitary in platypus (Ornithorhynchus anatinus) and short-beaked echidna (Tachyglossus aculeatus). J. Anat. , 221, 9-20. PMID: 22512474 DOI.
- ↑ Weisbecker V. (2011). Monotreme ossification sequences and the riddle of mammalian skeletal development. Evolution , 65, 1323-35. PMID: 21521190 DOI.
- ↑ Lefèvre CM, Sharp JA & Nicholas KR. (2009). Characterisation of monotreme caseins reveals lineage-specific expansion of an ancestral casein locus in mammals. Reprod. Fertil. Dev. , 21, 1015-27. PMID: 19874726 DOI.
- ↑ Ashwell KW. (2006). Cyto- and chemoarchitecture of the monotreme olfactory tubercle. Brain Behav. Evol. , 67, 85-102. PMID: 16244467 DOI.
- ↑ Ashwell KW, Hardman CD & Paxinos G. (2006). Cyto- and chemoarchitecture of the sensory trigeminal nuclei of the echidna, platypus and rat. J. Chem. Neuroanat. , 31, 81-107. PMID: 16198535 DOI.
- ↑ Semon R. Zur Entwickelungsgeschichte der Monotremen. Denkschriften der Medizinisch-Naturwissenschaftlichen Gesellschaft zu Jena (Embryology of the monotremes. Proceedings of the Medical and Natural Sciences Society in Jena). (1894) 5: 61–74.
- ↑ Rowe T, Rich TH, Vickers-Rich P, Springer M & Woodburne MO. (2008). The oldest platypus and its bearing on divergence timing of the platypus and echidna clades. Proc. Natl. Acad. Sci. U.S.A. , 105, 1238-42. PMID: 18216270 DOI.
Reviews
Johnston S. (2019). Challenges associated with the development and transfer of assisted breeding technology in marsupials and monotremes: lessons from the koala, wombat and short-beaked echidna. Reprod. Fertil. Dev. , 31, 1305-1314. PMID: 30991015 DOI.
Rowe MJ, Mahns DA, Bohringer RC, Ashwell KW & Sahai V. (2003). Tactile neural mechanisms in monotremes. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. , 136, 883-93. PMID: 14667851
Musser AM. (2003). Review of the monotreme fossil record and comparison of palaeontological and molecular data. Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. , 136, 927-42. PMID: 14667856
Belov K & Hellman L. (2003). Immunoglobulin genetics of Ornithorhynchus anatinus (platypus) and Tachyglossus aculeatus (short-beaked echidna). Comp. Biochem. Physiol., Part A Mol. Integr. Physiol. , 136, 811-9. PMID: 14667846
Temple-Smith P & Grant T. (2001). Uncertain breeding: a short history of reproduction in monotremes. Reprod. Fertil. Dev. , 13, 487-97. PMID: 11999298
Articles
Ashwell KW & Shulruf B. (2014). Vestibular development in marsupials and monotremes. J. Anat. , 224, 447-58. PMID: 24298911 DOI.
Selwood L & Johnson MH. (2006). Trophoblast and hypoblast in the monotreme, marsupial and eutherian mammal: evolution and origins. Bioessays , 28, 128-45. PMID: 16435291 DOI.
Ashwell KW. (2006). Cyto- and chemoarchitecture of the monotreme olfactory tubercle. Brain Behav. Evol. , 67, 85-102. PMID: 16244467 DOI.
Thorp BH & Dixon JM. (1991). Cartilaginous bone extremities of growing monotremes appear unique. Anat. Rec. , 229, 447-52. PMID: 2048749 DOI.
Keast JR. (1993). Innervation of the monotreme gastrointestinal tract: a study of peptide and catecholamine distribution. J. Comp. Neurol. , 334, 228-40. PMID: 8103529 DOI.
Manger PR, Fahringer HM, Pettigrew JD & Siegel JM. (2002). The distribution and morphological characteristics of cholinergic cells in the brain of monotremes as revealed by ChAT immunohistochemistry. Brain Behav. Evol. , 60, 275-97. PMID: 12476054 DOI.
Djakiew D & Jones RC. (1983). Sperm maturation, fluid transport, and secretion and absorption of protein in the epididymis of the echidna, Tachyglossus aculeatus. J. Reprod. Fertil. , 68, 445-56. PMID: 6864661
Semon R. Zur Entwickelungsgeschichte der Monotremen. Denkschriften der Medizinisch-Naturwissenschaftlichen Gesellschaft zu Jena (Embryology of the monotremes. Proceedings of the Medical and Natural Sciences Society in Jena). (1894) 5: 61–74.
Books
The Echidna: Australia's Enigma (Hardcover, 1999), by Peggy Rismiller (Amazon Link) "The oldest surviving mammal on the planet is also one of the most intriguing. Peggy Rismiller, the world's foremost echidna expert, traces the history of this fascinating animal that is native to Australia and New Guinea. A combination of mammal, reptile, and marsupial, echidnas produce milk, but unlike mammals, they are egg-laying creatures and, like marsupials, they have a modified pouch for nurturing their young. This odd animal has two backward-facing appendages and two forward-facing ones. These and other bizarre biological traits are discussed in detail in this thorough guide. Amazing photographs of echidnas enliven Rismiller's text, which includes Aboriginal tribal legends about the animal as well as the latest information on biological research being conducted today. With fossils dating back 120 million years, the echidna lived alongside dinosaurs, but unlike the giant reptiles, it survived. Its story and biology teach a fascinating lesson about endurance, survival, and sustainability."
American Museum Novitates (American Museum of Natural History) Van Deusen, H. M., and G. G. George. Results of the Archbold Expeditions. No. 90. Notes on the echidnas (Mammalia: Tachyglossidae) of New Guinea. American Museum Novitates, 2383:1-23 (1969)
Search PubMed
Search Jan2006 "Echidna development" 303 reference articles of which 20 were reviews.
Search PubMed: Echidna development | monotreme development
External Links
External Links Notice - The dynamic nature of the internet may mean that some of these listed links may no longer function. If the link no longer works search the web with the link text or name. Links to any external commercial sites are provided for information purposes only and should never be considered an endorsement. UNSW Embryology is provided as an educational resource with no clinical information or commercial affiliation.
- Pelican Lagoon Research & Wildlife Centre Echidna Research
- International Union for Conservation of Nature and Natural Resources Red List | Zaglossus bruijni Australasian Marsupial & Monotreme Specialist Group 1996. Zaglossus bruijni. In: IUCN 2004. 2004 IUCN Red List of Threatened Species. www.iucnredlist.org. Downloaded on 13 February 2006.
- The Australian Mammal Society Species Short-beaked Echidna | Long-beaked Echidna
- Access Excellence The National Health Museum (USA) Australian Mammals: Evolutionary Development as a Result of Geographic Isolation
- Science Alert CRCA Media Release 05/29 Echidna milk to reveal its secrets for dairy
- Echidna Gallery
- Wombaroo Food Products Echidna Milk Replacer
- Comparative Mammalian Brain Collections Echidna Brain Atlas
| Animal Development: axolotl | bat | cat | chicken | cow | dog | dolphin | echidna | fly | frog | goat | grasshopper | guinea pig | hamster | horse | kangaroo | koala | lizard | medaka | mouse | opossum | pig | platypus | rabbit | rat | salamander | sea squirt | sea urchin | sheep | worm | zebrafish | life cycles | development timetable | development models | K12 |
Glossary Links
- Glossary: A | B | C | D | E | F | G | H | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | Numbers | Symbols | Term Link
Cite this page: Hill, M.A. (2024, April 26) Embryology Echidna Development. Retrieved from https://embryology.med.unsw.edu.au/embryology/index.php/Echidna_Development
- © Dr Mark Hill 2024, UNSW Embryology ISBN: 978 0 7334 2609 4 - UNSW CRICOS Provider Code No. 00098G